Biology Concepts – innate immunity, acquired immunity, memory response, influenza
Your body is exposed to tens of thousands of foreign molecules every day. Some can do you harm, some can’t. Your immune system sorts them by matching receptors on immune cells to molecules on the foreign objects.
 |
Legos and biology are a good fit. They can be used to analogize the rearrangement T cell receptor genes or hypervariable regions of antibody genes, or they can be used to model the entire body. One scientist uses them to model building complex systems from repetitive units. And they’re fun. |
Thinkof the receptors as Legos; your DNA provides for several different types of Lego blocks to be made, and your immune cells can rearrange the different types and put them together as a receptor, so there can be millions of different receptors. Each immune cell has just one type of Lego receptor, although it may have many copies of that one form. Each different Lego receptor will fit, key in lock style, with a specific foreign molecule.
The receptors exist on many types of cells, and antibodies sometimes function as receptors when attached to the surface of specialized immune cells. Even circulating antibodies (Ab) in the blood take the form of key and lock systems, whether as single Ab, dimers (2) or pentamer (5) complexes.
The receptors exist on many types of cells, and antibodies sometimes function as receptors when attached to the surface of specialized immune cells. Even circulating antibodies (Ab) in the blood take the form of key and lock systems, whether as single Ab, dimers (2) or pentamer (5) complexes.
The immune system of higher animals can be described as several sets of pairs. Each member of a pair attacks a problem in a certain way, and has independent pathways, but each pair also has overlap and must work together in an overall response. We could spend weeks just on this system, but lets look at the major parts by describing each pair, from largest to smallest.
Innate immunityvs. adaptive immunity– the innate immune responses are fast but short. They don’t depend on your immune system recognizing the specific foreign molecule (antigen) with a specific receptor, but respond with the same types of reactions no matter what it is. Almost all plants and animals have some form of innate immune system.
Vertebrates take the immune system further. They have developed an adaptive immune system that does depend on your immune system recognizing the specific foreign invader. It then generates a tailored response to that one foreign organism or molecule. The faster, but more general, innate response helps the slower, but longer lasting and more specific, adaptive response to kick in.
 |
These are cartoons of an antibody. The model on the left is a much more realistic image. The Fc portion is the same through most antibodies (c= constant), while the gene rearrangement takes place in the light chain and heavy chain variable regions. The different variable regions are the Lego blocks that can be put together differently to make the millions of different antigen binding sites. |
Humoral immunityvs. cellular immunity– when an antigen is recognized by an adaptive immune cell (often through antigen presentation by the innate system), an early response is for the cell to divide and make more of itself. You don’t get sick from one bacterium infecting you; many infect you at once and then divide to become many more. You need many copies of that specific immune cell in order to battle the invading horde of bacteria.
The immune cells can generate an antibody response (humoral immunity) and/or trigger specific killing and directing cells to be produced (cellular immunity). The antibody (produced by B lymphocytes) is a protein that recognizes the specific antigen. The cellular immune response is mediated primarily by T lymphocytes.
However, B cell-produced antibodies are important for T cells to do their work, and antibodies also help the innate immune response to keep working after specific recognition has been made. In addition, the cellular immune response can control and ramp-up the humoral response. You see what I mean about each pair being separate but connected.
Effector T cellsvs. regulatory T cells– There are pairs of T cells as well. I use the term “effector T” cells to lump CD8+and CD4+ lymphocytes together (CD = cluster of differentiation markers on the cell surfaces). Effector T lymphocytes are either directly cytotoxic (CD8+, cyto = cell and toxic = damaging) or command (CD4+) the many adaptive responses. Effector cells are contrasted with regulatory cells, which include regulatory and suppressor T lymphocytes. The purpose of these cells is to stem the effector response so it doesn’t get out of hand; parts of the immune response are inflammation and non-specific cell killing – too much of that and you die too.
Memory Immune System– This last part of the immune response is not a member of a pair. When your innate immune system is activated, it ramps up, does its job, and hopefully is turned back off. The adaptive immune system responds to the antigen by producing more cells, antibodies and chemical signals (cytokines), and after the invader is vanquished you want this response to diminish as well. The innate system always starts over from zero, but the adaptive system remembers the infection you had.
 |
The dendritc cell on the left is an innate immune cell that works to present the antigen to the adaptive immune cells (Th1, Th2, and B cells). The adaptive cells reproduce and make cytokines to stimulate other immune cells. They also generate some memory cells that recognize the same antigen, but stay around for a long time and can react strongly and quickly. |
Duringthe adaptive response, some of the produced immune cells become “memory cells,” they still recognize the antigen from the initial infection, but hang around in larger numbers; in many cases they circulate in your body for the rest of your life. If your body sees that specific antigen again, the memory response can be re-initaited very quickly and very aggressively. You might be infected again, but your memory response is so fast and effective that you never know it.
In a world without vaccines, you are infected, get the disease, recover (hopefully), and then have a memory immune system for that antigen. Vaccines take the initial infection and disease out of the equation; you get to develop a memory without having had the experience!
As we discussed last week with smallpox, vaccines present your immune system with the antigen in the form of a dead or weakened pathogen, or just the antigen molecule itself. Your body doesn’t know the difference, it develops an adaptive and memory response just as if it were the real infection.
In the majority of cases, you develop memory B and T lymphocytes when infected or vaccinated. However, there are exceptions. Most antigens cannot fully activate B cells to make antibody, they have to be helped along by antigen-activated T cells. But there are T cell-independent antigens that can fully activate B cells on their own. In these infections, you can develop a B cell memory without a T cell memory.
On the other hand, there are other infections that develop a full memory response, but it is not useful. Influenza is an example of this. Influenza has been around for thousands of years; some years we have severe epidemics or even world-wide pandemics. The 1918-1919 Spanish flu pandemic killed over 50 million people, many more than the contemporaneous WWI (16 million deaths).
Flu is difficult to vaccinate against because it keeps changing. Influenza virus has two antigens, called H (hemagglutinin) and N (neuraminidase). These are the molecules on the virus particle that your body mounts an immune response against.
The H molecule on the viral coat binds to sialic acid receptors on respiratory cells and allows the virus to enter. When the newly produced viruses bud off of the cell, they place H on the cell surface, but there are still host sialic acid receptors there as well. These receptors would bind up the H and prevent the new viral particles from attaching to and infecting other cells, so the N molecule cleaves the sialic acid receptors from the new viral particles.
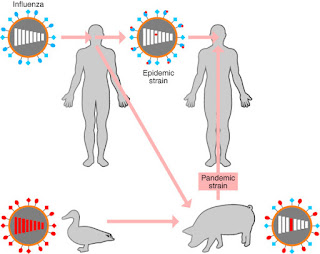 |
Influenza virus can mutate by antigenic drift or antigenic shift. The top line shows that by passing from person to person, the antigens (and virulence) shift slightly. The lower line shows that by passing through other animals and recombining, the antigens can have small or big changes. When shifted virus moves into humans, it’s a recipe for a pandemic. |
Theproblem arises when the H and N antigens mutate.... and they do. Scientists have identified 16 different classes of H’s and 9 different N’s, and they can be paired up in many combinations. Small changes (antigenic drift) usually mean that memory might have a slight protective effect, and major epidemics do not occur. But major changes in H and N (antigenic shift) mean that previously infected people have no memory protection.
Different strains of influenza virus can infect the same animal (often pigs and ducks – thus avian flus and swine flus) and can mix their H’s and N’s. What emerges and might be transmitted to humans can be a virus with H’s and N’s similar to years past, or with new H’s or N’s. That is why a new vaccine must be produced each year, after scientists see which H’s and N’s the new virus has and how much they have drifted. Avian flu is H5N1, while swine flu is H1N1. However, antigenic drift means that each H1N1 will not be exactly like the previous H1N1 to emerge. The 1918 pandemic was caused by an antigenically shifted H1N1 sub-strain.
Like flu, other infections may not provide life-long memory. If the memory response is weak or the initial response was not strong, then memory may fade over time. This is why some vaccinations require boosters in later years. A fading of the memory response to influenza is also implicated in the need for yearly vaccinations.
 |
Here's a great book that discusses both the biology and sociology of influenza. There are great personal stories as well as medical detective work. This pandemic was a jolt that brought infectious disease research into a new century. I highly recommend it. |
Now for the exception to the exception. Influenza changes each year, so memory does not help much, but a 2010 report from scientists in Hong Kong suggests that prior exposure to any seasonal influenza (either by infection or vaccination) might have been a contributing factor as to why the 2009 pandemic of antigenically shifted swine flu (H1N1) was much milder than expected.
The 2009 seasonal flu vaccine did not have any cross-reactivity with pandemic H1N1, so the scientists suggest that previous years seasonal influenzas did generate some memory response that was partially effective against 2009’s H1N1 swine flu. Cross-reactivity means that the H and N antigens were not identical to previous version; the Legos don’t fit together exactly, but they were similar enough to fit together and initiate a partial response. Once again, we see that getting sick may save your life down the line.
Next week will look at examples wherein having one disease can protect you from catching another.
For more information or classroom activities, see:
innate immunity:
adaptive immunity:
memory immune response:
influenza virus:
http://www.xvivo.net/zirus-antivirotics-condensed/