Biology concepts – flagella, bacteria, prokaryotes, eukaryotes, undulipodia, axoneme, basal body, centriole
You find yourself transported back to sixth grade grammar class. You barely fit in the desk and your clothes are out of style.... again. You don’t know how you got there, but the immediate problem is that Mrs. Belcher has just called your name to answer the next question. What are homonyms?! You stare back at her with terror in your eyes.
The axoneme (axo = axis, and neme = thread) of the undulipodia has a very distinct structure which is best appreciated when you look at it in cross section. The long microtubles appear as small circles in the cross cut, but they are arranged very precisely (see picture). They come in doublets, and there are nine sets of them surrounding a central doublet (called 9 + 2 or 9(2) + 2). This is very different from the single hollow tube of the prokaryotic flagellum.
The motions the two different mechanisms produce is different too (a homophone). While the prokaryotic flagellum spins like a propeller, the sliding of the microtubules makes the undulipodium wave back and forth, like a field of wheat on a breezy day, or how a snake might thrash if you held his head down (don’t just hold his tail down, he’ll bite you).
Our two systems still look similar, but we see how they are not so similar in structure and action. Since eukaryotic cells are so much bigger than prokaryotes, the first flagella to be examined and found to have the 9(2) + 2 structure were in sperm tails.
A couple of parasitic protozoans that live in bristle worm guts show differences in their axonemes. Lecudina tuzetae sperm tails have a 6(2) + 0 structure, while the Diplauxis hatti protozoan sperm has a 3(2) + 0 axoneme. Described in a 1980 paper, this is the simplest motile undulipodium known – as of now.
Both eukaryotic flagella and cilia have the microtubule and dynein arm structure, with centrioles for their basal bodies. There are exceptions to the cilia that we will look at in a few weeks, but the biggest difference between them, besides their length, is that cilia occur in groups, while flagella are usually found in ones or twos. You could say that cilia and flagella are like synonyms, they have almost the same meaning (and structure), although they are two different things.
Prensier, G., Vivier, E., Goldstein, S., & Schrevel, J. (1980). Motile flagellum with a "3 + 0" ultrastructure Science, 207 (4438), 1493-1494 DOI: 10.1126/science.7189065
Idei, M., Osada, K., Sato, S., Nakayama, T., Nagumo, T., & Mann, D. (2012). Sperm ultrastructure in the diatoms Melosira and Thalassiosira and the significance of the 9 + 0 configuration Protoplasma, 250 (4), 833-850 DOI: 10.1007/s00709-012-0465-8
Feistel K, & Blum M (2006). Three types of cilia including a novel 9+4 axoneme on the notochordal plate of the rabbit embryo. Developmental dynamics : an official publication of the American Association of Anatomists, 235 (12), 3348-58 PMID: 17061268
 |
Everyone has the dream where you show up for a class that you didn’t know was on your schedule, only to be having a test. But in second place is the dream where you are back in elementary school, or maybe the principal’s office. Above is a picture of every teacher I had in elementary school. |
But your study of word roots may help you survive. Homo- means same, while -nym means word. OK, it’s coming back to you. Homonyms can be words that are spelled but have different meanings and origins (called homographs) or words that are pronounced the same but have different spellings and meanings (called homophones). Yes! The class cheers, and Mrs. Belcher is more than mildly surprised. Crisis averted.
I have no idea how you got transported back to grammar school, but your question and answer is very timely to our discussion today. Homographs, like minute (min-it, a short time) and minute (my-noot, a small amount), look the same, but have different meanings. Homophones, like to,too, and two sound the same but mean different things and have different origins. This is very much like the differences in prokaryotic flagella and eukaryotic undulipodia. Too much of a stretch for you... maybe.
We have seen that bacterial flagella are long, whip-like structures protruding from the cell that can aid in motility. So are eukaryotic undulipodia. They look very similar, yet we are going to see they have very different structures, mechanisms of function, and origins – just like homophone and homonyms. So maybe the analogy wasn’t so far off.
Both flagella and undulipodia extend from the cell surface with a long tail. But in the prokaryote, this was made of small subunits of flagellin proteins. In the undulipodia, the structure is called the axoneme, and is made of long microtubules of tubulin protein. Already we have significant differences between two things that look very similar.
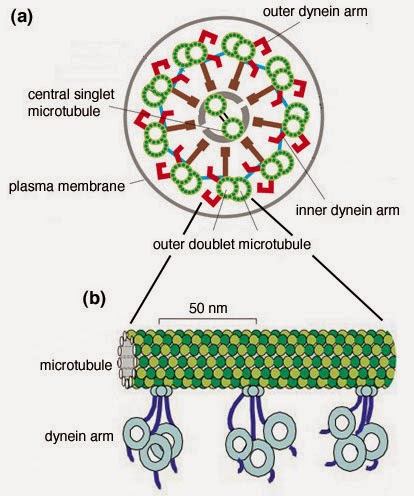 |
A cross section of an undulipodium axoneme looks like this – although I’m not sure they’re color coded in the cell. Notice how the inner are connects the nested tubule of one doublet to the outside tubule of the next doublet. The inner arms are responsible for the degree of bend, the outer arms are involved in the rate of movement. There are other proteins involved, but we aren’t getting that detailed. Maybe you want to do that on your own. |
The way the axoneme is built is also the key to how it works. The different microtubule doublets are cross-linked by protein complexes called dynein arms. There are inner arms and outer arms. An inner arm connects one microtubule from one doublet to another microtubule of the adjacent doublet. When one doublet slides further out from the cell body and the connected adjacent doublet doesn’t, this creates stress and the whole thing must bend to maintain the connections.
So, walking proteins are how the undulipodia create their whip-like action. There is a walking system analogous to dynein arm movement that has developed in many animals. We have looked at muscle contraction before. Just like myosin heads walking along actin filaments that are anchored to the muscle cell membrane, the dynein arms on some doublets start to crawl up or down the adjacent microtubule of an undulipodia creating a bend and then they can reverse to whip back the other direction. This is very different from the spinning motor of the bacterial flagellum.
 |
Pay attention to these cartoons, they show how the undulipodia bend. ATP powers the sliding of the dynein arms. As they move down one tubule, that filament moves up. If the filaments are anchored in the membrane, as they are in undulipodia, the movement creates tension and a bend. The cartoon on the right is a good summary. |
The undulipodium motion often occurs in just one plane, back and forth instead of all around, but that doesn’t mean it has too limit itself to that. It can spin too; it just takes a very coordinated sliding back and forth of microtubules.
Another difference between prokaryotic flagella and eukaryotic undulipodia is in how they are powered. We saw that flagella in bacteria get their force from the spinning motor, and the motor gets its energy from an ion gradient across the membrane. But in eukaryotes, it was seen early on that if you strip the membrane off of an undulipodium, and added ATP, they start to move.
Yes, all undulipodia are held within the membrane. Some bacterial flagella are membrane covered, but all eukaryotic versions are sheathed in plasma membrane. But back to the ATP. Exposing the naked undulipodium to ATP, even on a dead cell, can initiate the dynein walking and microtubule sliding, so it is definitely ATP powered.
Also, this points out that the power for the bacteria movement comes from the motor in the base, but the eukaryotic movement is in the axoneme, not the base. The basal body that anchors the eukaryotic undulipodium into the cell membrane is amazing in its own way. The basal body is actually a centriole, the same structure that helps to move the chromosomes apart in the spindle apparatus during mitosis. We’ll come back to this double duty organelle in a a few weeks.
 |
Chlamydomonas algae species have a double flagella for swimming. A single stroke as shown above. The power strokes on top is followed by the recovery stroke below. Put all the numbers together and looks like someone swimming underwater. The picture of the organism is there because it’s always better to see the real thing as compared to a cartoon. |
But soon after that, the protozoans were discovered to have undulipodia as well. Organisms from the algae genus Chlamydomonas have two long undulipodia that they use for motility. Located at the front of the cell, their movement pulls the alga through the water. But protozoans are just as likely to have undulipodia that push them through water. They can work both ways.
Amazingly, when a Chlamydomonas finds itself out of water, the undulipodia resorb in short order. Nature hates to waste energy, so why maintain a boat motor if you’re not in the water. But place them back in a liquid environment and the two structures will reassemble with in an hour – with the same structure 9(2) + 2 and working the same exact way.
Of course, saying they all have the same 9(2) + 2 structure is an invitation to find exceptions, and science has found them. A 2006 studyfound that rabbits are quite the rule breakers. Sure, they have 9(2) + 2 axonemes, but they don’t stop there. Some rabbit embryo undulipodia show a 9(2) + 0 structure, where the central doublet is completely missing, yet the structure functions just as other motile undulipodia.
What’s more, rabbits can also have 9(2) + 4 axonemes, with double the number of central microtubules. Again, they function just fine. Is there a reason for these variations – maybe, but maybe they are just mutations that didn’t have a negative impact, so they were retained.
 |
You know how annoying it is when you touch a cactus and those little bristles get stuck in your skin? Well, don’t touch a bristleworm. They’re lined with those painful bristles – hence the name. Somebody studied these worms, and found out they have parasitic protozoans in their gut. Then someone studied those parasites and found that they have sperm. And someone studied the sperm and found that they have unique axoneme structures. I love science. |
The undulipodium basal body (born as a centriole) can have exceptions as well. The vast majority have a structure of 9(3) + 0, where instead of doublets, they have triplets. This makes sense since they need to be strong to support the axoneme.
But diatoms, very small algae cells protected by a silica shell, can have sperm that look very different, according to a 2013 study. Their basal bodies have been observed to have doublet microtubules, and are very similar to the axoneme. Even weirder, a couple of insects feel the need to go big with their basal bodies. Acerentomonon microrhinus, a primitive hexapod insect has sperm tail basal bodies with 14 microtubule doublets, while Sciara coprophila, a fungus gnat (see picture), has up to 90 singlet microtubules in its sperm basal body.
We have talked a lot about sperm tails and protozoan motility structures, and these undulipodia look the most like bacterial flagella. But undulipodia come in a couple of flavors; those longer than 40 m or so are called flagella while the shorter ones are called cilia. See the naming problem and why Lynn Margulis came up with undulipodia?
 |
This is a fungus gnat – it sounds like they eat fungus, but nope. Only the larval form feeds; the adults never eat. That’s OK, they don’t live very long at all. They overwinter in their adult form because they are unique in that they can both tolerate freezing weather and freeze themselves without damage. It must be important, since their sperm tails are loaded with 90 microtubules in the basal body – everyone knows that massive numbers of microtubules is the best way to avoid cold damage. |
Next week – Halloween is coming, so what better time to have a discussion of genetically modified foods and an 19th century teenage girl who wrote the best science fiction book ever.
For more information or classroom activities, see:
Undulipodia –