Biological concepts – primary cilia, sterocilia, kinocilium, Usher syndrome, actin, microtubule, signal transduction, sensory receptor, mechanoreceptor
 |
The USS Oriskany (above) was scuttled in 2006 to create an artificial reef off of Pensacola Florida. In 2012, the US government effectively ended its policy of creating artificial reefs this way because of concern for leaking toxins from the ships to the marine life. But is was a good way to find a new job for something broken. |
Something that seemed broken because it couldn’t move was given an important new job that didn’t require motility. Remember that analogy as we talk about today’s subject in cilia. Although the order might be reversed.
We spoke last week about how nematodes are the only animals that don’t have cilia. Eukaryotic cilia and flagella (together, the undulipodia) are organelles that move, and in turn may move cells. It turns out that cilia have some exceptions – some don’t beat, and some can’t move at all - so what good are they?
Motile cilia, the kind we have been talking about for the past couple of weeks, are also called 2˚ cilia. If there are 2˚ cilia, I think that pretty much implies that there must 1˚ cilia– and they’re what we will talk about today.
Primary cilia, while less well known, are found on many more cell types than are motile cilia. Motile cilia in mammals are located on male gametes (as flagella), on respiratory epithelium of the lower and upper respiratory tract, fallopian tubes near the ovary and epididymal cells of the testes, and the ependymal cells lining the ventricles of the brain.
Primary cilia are apparent on cells of most types, when they are quiescent (just hanging out, doing it's job). If the cell re-enters the cell cycle and starts to divide or differentiate, the primary cilium will resorb and then reappear in daughter cells once they become quiescent.
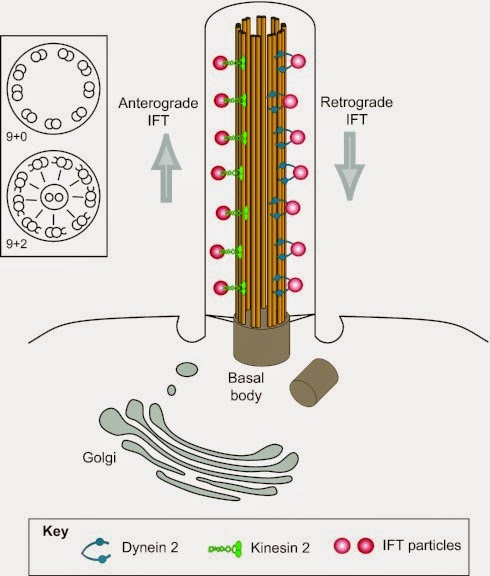 |
Primary cilia have basal bodies and IFT, but their microtubule structure is different. They don’t have the inner singlet microtubules, so the outside ones can’t slide past one another. They do have outer dynein arms, and those are important for retrograde IFT. See below, kinocilium don’t even have the outer arms. |
Another exception with primary cilia is that their microtubule axoneme can change as it goes out to the end of the cilia. It may start out as 9(2) + 0, but at the distal (far) end it's 9 + 0 in nematodes, algae, and in the nose, pancreas and kidneys of vertebrates. All of those count as exceptions too!
In addition, primary cilia are of differing lengths, but most are much shorter than motile cilia. Some don’t even extend from the surface of the cell membrane. However, they're built by IFT (intraflagellar transport) just as 2˚ cilia are, and IFT is important for their functions as well.
So, can a broken cilium have a specific job? If they don’t beat to move a cell or the environment around the cell, then what do primary cilia do? The answer is - just about everything. Primary cilia serve as mechanoreceptors, chemoreceptors, photoreceptors, as well as osmolarity, temperature, or gravity receptors. Think of primary cilia like weather balloons. They stick out into the environment and probe the conditions in the area. They send the data back and the cell can act on it.
As mechanoreceptors, primary cilia might not beat, but they can be moved. They bend in response to flow across the surface and the bend brings a pivot at the level of the basal body – yes, primary cilia have basal bodies just as motile cilia do.
 |
Kidney cells that line the tubules have primary cilia to sense the urine flow. The cilium is bent, and this triggers a change in calcium influx. The change is then transferred to the adjacent cell via calcium channels that cross both membranes. |
Primary cilia have an asymmetry so that they recognize right from left. In the kidney, the flow is based on orientation, all primary cilia bend in the same direction, toward the anterior. The anterior bend signals for increased calcium influx and then this signal is transmitted to adjacent cells. The uniform gradient (a-p) works cell to cell, and this leads to consistent a-p orientation of the mitotic spindle (which also uses basal bodies in the form of centrioles). The result is that the progeny cells of dividing renal epithelium have the same orientation as the parent they replace.
Back to our nematodes from last week. Primary cilia are the only cilia roundworms have. C. elegans, the roundworm that is used as a laboratory model, is made up of exactly 959 cells – exactly. Sixty of those cells, all sensory neurons, have primary cilia that stick out into the environment via pores called sensillae.
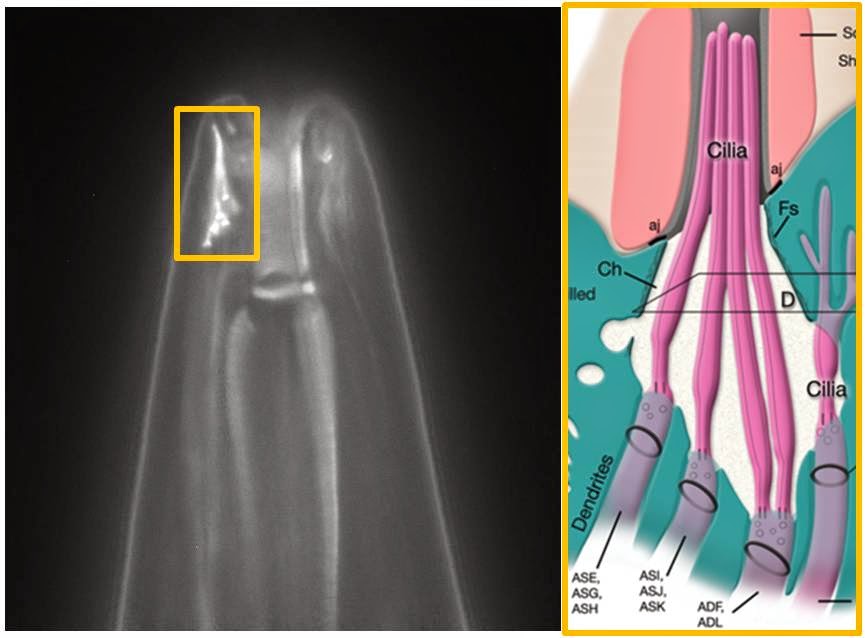 |
The left photomicrograph has labeled dendrites for sensory neurons in C. elegans. The right cartoon shows how the primary cilia from these neurons stick into the pore that then helps them sense the environment around the roundworm. |
It’s through the interaction of these primary cilia with the worm's immediate environment that it senses its world. This is what passes for a roundworm brain – but your brain has them as well. Especially in the retina of your eyes.
The photoreceptors that absorb light energy and transfer it to electrical impulses are located on a single primary cilium on each retinal cell. The axoneme is used to move photosensitive pigments (like retinal in rhodopsin, see below) back and forth from the receptor to the cytoplasm.
Primary cilia also act as chemoreceptors. In brain proper, they work in formation of new memories – mice without primary cilia can’t remember new objects or recognize objects they have already learned. They can remember the location of the object just fine, just not the object itself. We will talk about primary cilia in the brain much more next week.
Now we can take this discussion a couple of steps further to talk about two ciliary exceptions. There are nonmotile 1˚ cilia, motile 2˚ cilia, and then a third structure called a kinocilium. From the Greek for moving eyelash, the kinocilium is poorly named. Described in guinea pigs in 1989, they don’t move like a blinking eyelash or even like a motile cilium; they lack the inner dynein arms and central microtubules that would allow them to be motile. But, they can move horizontally across a cell surface.
 |
As this movie travels down the photoreceptor, notice the vertical basal body/axoneme on the left. This is a primary cilium! The microtubules help move photo pigments up and down the cilium. |
If hair cell kinocilia are poorly named, then hair cell stereocilia are down right liars. They aren’t cilia at all. The characteristics of cilia include that they are microtubule extensions of a basal body modified from a centriole. They may be motile or nonmotile, but their functions are mediated by moving signaling, structural, or receptor molecules up and down via intraflagellar transport proteins.
None of that applies to sterocilia! They're built from actin not microtubules. They do not have an intraflagellar transport system. They have no basal body. They are very similar to the microvilli of your gut epithelium, but nothing like cilia, except for the fact that they stick up from a cell.
The hair cells work by using the sterocilia as mechanoreceptors. In the cochlea, they bend in response to fluid movement based on vibrations of sound. In the semicircular canals, they bend in response fluid movement as a result of changes in head position. When the sterocilia bend, it generates an action potential in neurons that go to the brain.
 |
Hair cells of the cochlea can be damaged by loud noise. The left images are the normal (top) hair cell sterocilia, and the same sterocilia after a loud noise (bottom). The right images show a series of hair cells in normal condition, and after a long time exposed to loud noise. Turn down your music – do you think the hair cells in the damaged cochlea work well? |
The sterocilia start to grow longer, with the ones closest to the kinocilium being the longest. They line up to look like a choir on risers in front of the taller kinocilium. Now they are ready to function. At this point the kinocilium disappears! If you look at working hair cells, you won't find the structure that mediated their development.
So we have two new ciliary structures - neither of which act like cilia. That’s weird enough, but it gets weirder. There is a disease that affects both hearing and vision because it messes with the primary cilia of the retina and the sterocilia of the ear. But we just learned that those are two completely different structures!
 |
People often use "tunnel vision" to explain the field changes in retinitis pigmentosa, the kind of progressive blindness in Usher syndrome. But really, it’s more like backing into a tunnel, one that never reaches the other side. Think of running this clip backwards. |
As we said last week, nature hates a unitasker. There are at least 11 proteins that work in development and working of both sterocilia AND primary cilia, even though they look and are built completely different. A mutation in one of those proteins affects all three systems. Maybe it would be better if sometimes a protein had just one job, fewer things could get screwed up if something goes wrong with it. Would you rather have reduced vision, reduced hearing, bad balance, or all three?
All this knowledge leaves us with an unanswered question - did the sensory primary cilia develop from motile cilia, or did motile cilia develop from the primary version? Did broken motile cilia develop a new job, or did 1˚ cilia learn how to dance after they had learned their first function? Hmmmm.
We've barely touched the functions of cilia that don’t even move. In the next couple of weeks, we will see how primary cilia keep you from being fat, and how they will be crucial for long-term space travel. Then we can figure out how they give you a right and left hand.
We've barely touched the functions of cilia that don’t even move. In the next couple of weeks, we will see how primary cilia keep you from being fat, and how they will be crucial for long-term space travel. Then we can figure out how they give you a right and left hand.
For more information or classroom activities, see:
Primary cilia –
Hair cells –
http://education-portal.com/academy/lesson/the-ear-hair-cells-organ-of-corti-the-auditory-nerve.html
Visual photoreceptors –
Usher syndrome -