Biology concepts – botany, monoecious, dichogamy, imperfect and perfect flowers, self-pollination, cross-pollination, self-incompatibility, heterostyly
There are a few ways for animals to make new animals. Asexual reproduction is possible in a few species, while sexual reproduction is much more common. In between, there are the hermaphrodites. These animals carry both sets of reproductive organs.
Apple trees are monoecious perfect, although most are pollinated by other apple trees, not themselves (see below). Interestingly, in order to get the most and best fruit, the king flower (the largest and first bloom of a cluster) must be pollinated and be pollinated first.
The small Spanish flower Silene littorea was recently (2013) found to be mostly gynomonoecious. Before this, it was thought that this species had two populations of plants, but the seed numbers and variable numbers of female flowers show that being gynomonoecious helps significantly in producing more seed with less flower investment.
A pollinator well-designed to gather pollen from a long anther would be poorly designed to accidentally (it’s almost always an accident) deliver that pollen to a short style. So it is unlikely that self-pollination will take place.
Both mechanisms involve genes that can rearrange to form many slightly different gene products. One individual will have the same rearrangement of the gene in its pollen and its egg. Pollen of one type will not work with an egg of the same type. It works in the exact opposite fashion as self-recognition proteins in humans. In that case, tissues with different HLA markers are attacked as foreign; in plants, pollen and egg of the same rearrangement will be shut down.
So these are ways to prevent self pollination in perfect and imperfect monoecious plants. But monoecy can get weird on its own in an attempt to prevent selfing:
One system of dichogamy comes about if the flowers of the monoecious plant are perfect. In this case, the structures are all there, but the timing for maturity is different. The flowers of Scyphiphora hydrophyllacea, a mangrove shrub, are perfect and protandrous, whileCenchrus clandestinus, a Hawaiian grass, has perfect, protogynous flowers. The flowers are structurally perfect, but functionally imperfect.
In perfect protandrous plants, the pollen matures and is carried away (ind, insects, animals, etc) before the ovules mature on the same flower - so no selfing. In perfect protogynous, the opposite is true. The early maturing eggs must procure pollen from individuals who have already had mature eggs and have changed to produce pollen.
The key is in the timing. Not all the individuals of a protogynous perfect population will flower exactly at the same time. So some will have moved on to being male while others are still female. This would then provide pollen for them without resorting to self-pollination.
Casimiro-Soriguer I, Buide ML, & Narbona E (2013). The roles of female and hermaphroditic flowers in the gynodioecious-gynomonoecious Silene littorea: insights into the phenology of sex expression. Plant biology (Stuttgart, Germany), 15 (6), 941-7 PMID: 23174011
Pérez-Barrales, R., Vargas, P., & Arroyo, J. (2006). New evidence for the Darwinian hypothesis of heterostyly: breeding systems and pollinators in Narcissus sect. Apodanthi New Phytologist, 171 (3), 553-567 DOI: 10.1111/j.1469-8137.2006.01819.x
Miller, J., & Diggle, P. (2003). Diversification of andromonoecy in Solanum section Lasiocarpa (Solanaceae): the roles of phenotypic plasticity and architecture American Journal of Botany, 90 (5), 707-715 DOI: 10.3732/ajb.90.5.707
 |
This clip shows the mating of hermaphroditic leopard slugs. Each may provide male gametes for the other, or it may just go one way. They hang from a branch to do this, and the male reproductive organs spiral around one another. The trait has gone mad – in some species, the male organ has reached 92 cm long! |
Some gastropod (snails and such) breeding is strictly sexual; they have male and female snails (marine), while most terrestrial snails are hermaphrodites. But even then, most need another gastropod to mate with. Each hermaphrodite fertilizes the eggs of the other. There are the rare cases where a hermaphroditic slug will self-fertilize and produce clones, but this is the exception, not the rule.
These examples show that weird reproduction does take place in animals, but the plants have us beat by a long shot. Even the simple types of flower breeding systems turn out to be not so simple. Sometimes being both sexes is the easy part.
If a single flowering plant (angiosperm) can produce both male and female reproductive cells (pollen and egg), the plant is called monoecious (one house). This represents a nice tight bundle, reproduction wise. One plant can make the male gamete cells (pollen) and the female gamete (egg) – it’s a hermaphrodite. Let’s look at the types of monoecious flowering plants, maybe it's not so simple:
Monoecious perfect - These plants have flowers that contain both the male reproductive structures and the females reproductive structures, so they are said to be perfect flowers. Because they occur on the same flower, any time the plant flowers, the blooms have both female and male structures and qualities. These are true hermaphrodites.
 |
The king flower of the apple blossom is the key to getting good apples. It has to be pollinated for the cluster to all produce apples. The king flower opens first, is pretty, and smells good. |
Monoecious imperfect– In the simplest form, monoecious imperfectplants have some male flowers and some female flowers, but they both bloom on the same plant. The flowers themselves aren’t hermaphroditic, but the plant is, since it has both male and female structures.
A single American chestnut tree will have both male and female flowers at the same time. Some long catkins (an arrangement of small flowers on a single stem) have only male flowers while others have male flowers at the tips and female flowers at the base.
Male and female flowers on separate individuals at the same time like with the chestnut is one form of monoecious imperfect, but there are others as well.
Gynomonoecious or andromonoecious – These are a mixing of perfect and imperfect flowers on the same plant. Gyno- means female, so these plants have imperfect female flowers AND perfect flowers at the same time. Andro- means male so, you guessed it, they have male flowers and perfect flowers.
A 2003 study of four Solanum (a large genus that includes potatoes and tomatoes) species showed that the number of male flowers compared to the number of hermaphroditic flowers can vary greatly. Some species were about 7% male flowers (weakly andromonoecious), while others were 69% male flowers (strongly andromonoecious). Weak species would change the number of male flowers produced according to how much fruit was produced, but strong species made the same number of male flowers no matter what.
 |
The American chestnut was a towering species until the late 1800’s when a nursery owner imported some Japanese chestnuts that had a fungal parasite. The American version had no immunity, and we lost these huge trees in short order. |
Problems with Monoecy- With either perfect flowers or imperfect flowers one a single plant, it’s possible for the pollen of an individual to fertilize an egg on the same individual – self-pollination. However, self-pollination isn’t always a good thing. Self-pollination produces clones of the parent that provides both the pollen and egg. Pollen from one plant fertilizing the egg of a second individual is called cross-pollination.
Cross-pollination promotes genetic diversity. Clones tend to build up genetic mistakes, while cross-pollination help to spread genes through the population and makes the species more likely to possess genes that might help them withstand changes in the environment. Therefore, many plants take steps to prevent self-pollination and promote cross-pollination.
Heterostyly (hetero = different, and style = part of the female reproductive organ) prevents “selfing” in many animal pollinated plants. In this case, a certain species will have two morphs (shapes) of flowers. One will have a long anther (pollen producer) and a short style (where the pollen is deposited and grows down to egg). The other will have short anthers and long styles (see picture and caption to below).
 |
Morph 1 and Morph 2 are the same species, but different individual plants. The large insect pollinator can easily get pollen from A anthers (right) and deliver it to A pistils (left), but how would it get pollen to the B pistil (right)? The reverse is true for the smaller pollinator. Therefore, Morph 1 can pollinate Morph 2 and vice versa, but neither can pollinate their own morph. This is heterostyly. |
However, that same pollinator would be well-designed to deliver its pollen load to a flower with a long style, the kind found on the other morph of individuals of the species. This would promote cross-pollination. The strategy is equally successful for those pollinators best prepared to gather pollen from short anthers.
It is not known whether the move to heterostyly in some plants has been driven by genetics to avoid inbreeding or by pollinators and the need for efficient fertilization. A 2006 study in Narcissus flowers looked at both genetics and pollinator efficiency in breeding and concluded that the pollinator driven evolution was supported to a greater degree. This agrees with the pollinator hypothesis that Darwin proposed almost 150 years ago. He was pretty smart.
In other cases, the position of the flowers may discourage self-pollination. For instance, some wind pollinated fir trees have female cones up high and male cones down low. The pollen from the male flowers might travel on the wind and gain altitude to fertilize female flowers on adjacent trees, but it is extremely unlikely that the pollen would be blown straight up to fertilize the female flowers of the same tree.
The most common mechanism to prevent selfing is self-incompatibility. There are two main mechanisms of self-incompatibility; they both work at the genetic level to make sure that the pollen of a particular individual will not successfully fertilize an egg of the same plant.
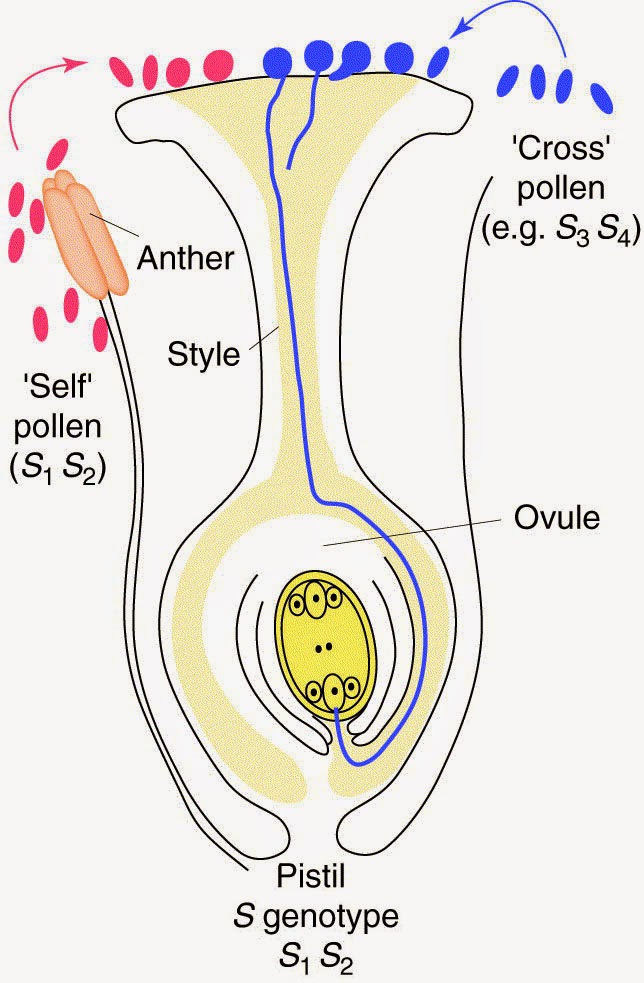 |
The blue pollen has one rearrangement of the compatibility genes (S3, S4), while the red pollen has a different rearrangement (S1, S2). If the ovule genome has S1, S2, then S1, S2 pollen landing on the stigma will be destroyed. S3, S4 pollen won’t be recognized and can grow pollen tubes to fertilize the S1, S2 egg. Self pollen is incompatible with the same egg; this promotes cross-pollination. |
So these are ways to prevent self pollination in perfect and imperfect monoecious plants. But monoecy can get weird on its own in an attempt to prevent selfing:
Dichogamous monoecy– This breeding system probably evolved as a way to prevent self-pollination in monoecious plants. The pollen and ovule mature at different times. This is equivalent to having a flower (plant) that can change its sex in just a short period of time, and these count as additional monoecious breeding schemes. Some animals can do this, but the change takes place over the period of a lifetime. Here were talking about in the period of a few hours.
If an individual plant can change its sex over a short period of time within one growing season, then it is called dichogamous (dicho = in two, apart, and gamous= gametes), also called sequential hermaphroditism or temporal dioecy. But which comes first male or female? If the plant first produces male flowers, then it is termed protandrous (proto = first). Protogynous is name for those that are female first. This is a great way to prevent self-fertilization and there are a couple of ways plants can employ dichogamy.
 |
This chart will help explain the different monoecious breeding systems. Each large circle is a population of plants of the species. The circle with cross means female flower, the circle with arrow means male flower, and the circle with both means perfect flower. The line arrow with a “t” means a change as time passes. For dichogamy, the flowers may be perfect or imperfect, but they function as male or female at each time point. |
In perfect protandrous plants, the pollen matures and is carried away (ind, insects, animals, etc) before the ovules mature on the same flower - so no selfing. In perfect protogynous, the opposite is true. The early maturing eggs must procure pollen from individuals who have already had mature eggs and have changed to produce pollen.
The other dichogamous possibility for monoecious plants is when they have imperfect flowers. This means that the plant would make flowers of one sex first, and then grow separate flowers of the other sex later on. The separate flowers are still on the same plant, but self-pollination isn’t possible because they aren’t there at the same time.
Corn is an imperfect, protandrous plant. This is why country kids detassel in the summer. The tassel is the male flower. If you remove it (de-tassel), it will prevent possible selfing when the female flowers come out. You can create hybrids by planting a few rows of a specific breed of male corn at the end of the rows.
It’s much harder to find an example of an imperfect, protogynous plant. In general, doesn’t it seem silly for a species to produce their female flowers first? They need pollen from the males in order to be fertilized, but if they’re all female, who provides the pollen? It doesn’t seem logical.
 |
The left image is the female flower of corn, every silk is a flower that can be pollinated. Each one that is will produce a corn kernel on the ear. The right image is the male flower, the tassel. This get pulled off by teenagers trying to make money for that prom dress or new stereo. |
But why no protogynous imperfectplants? The wasted energy in making purely female flowers very early when little pollen is present probably dooms this breeding system. At least with perfect protogynous, they get some benefit by dispersing pollen later from the same flower. No extra energy is consumed in producing an entirely different flower.
Duodichogamy - This system can help with the timing issue above; it’s dichogamy taken a bit further. Instead of being one sex then the other, they go back and forth and back and forth – like Mystique of the X-Men.In Bridelia retusa, a tropical tree from India, the switching between male and female occurs several times within a single week! To my mind, this is like sexual dimorphism in antlered males of some species - the antlers just keep getting bigger and bigger. Where will it all end - how many times will this plant change sex?
Next week - another mechanism for preventing self-pollination is to separate the reproductive parts to different plants. These are the dioecious plants.
For more information or classroom activities, see:
Monoecy –
self-pollination and cross pollination –