Biology Concepts – platyhelminthes, asymmetry, bilateral symmetry, evolution, cephalization, natural selection, fish, lepidophagy
The Great Barrier Reef off the northeast coast of Australia is alive. Reefs are made of the exoskeletons of coral polyps, with the new corals growing next to and on top of the older ones. With all the nooks and crannies available, coral reefs are some of the most diverse ecosystems on Earth, with thousands of species per square mile.
But we begin with a challenge –as we discuss the different animals that break symmetry in the next few posts, see if you can find something else that is common to many of them.
Different host species fish have different gill anatomy, so the attachment point and position will be different. The haptor(s) have to be located where attachment is possible. This means that they may be on one side of the body or the other, or two on one side and one on the other, etc.
Cichlid fish are one of the most diverse family of animals known, with more than 1700 known species. They are found in the Old World and the New. They exhibit some amazing adaptations, especially when many are found in one place, Lake Tanganyika on the border of Tanzania and the Democratic Republic of the Congo.
Mind you, this isn’t eating the scales of dead fish, or eating the scales that drop off live fish. Lepidophagy means eating the scales that are still attached to live fish. It’s a fish smorgasbord. This is both good and bad. Scales on live fish grow back fast, so there is always a ready supply of food.
The species P. microlepis has developed one particularly amazing way to help it eat the scales off of neighbor fish. It’s mouth and jaws have evolved so that they bend sideways. There are right-mouthed P. microlepis and left-mouthed individuals. The difference is obvious, right-mouthed individuals will feed only from the left side of their prey and vice versa. They have an asymmetry, or lateralization, of both anatomy and behavior
The cycle will begin again after left-mouthed individuals come to dominant in the population. Back and forth the population will go. If the population stayed 50/50, nobody would gain an advantage, and overall, they would all suffer. The system only works if a small number develop opposite to the majority. The gene regulatory complex that controls if an individual will be right- or left-mouthed must be very complex if it can take into account that a few need to be lateralized the other way.
Takeuchi, Y., Hori, M., & Oda, Y. (2012). Lateralized Kinematics of Predation Behavior in a Lake Tanganyika Scale-Eating Cichlid Fish PLoS ONE, 7 (1) DOI: 10.1371/journal.pone.0029272
Lee, H., Kusche, H., & Meyer, A. (2012). Handed Foraging Behavior in Scale-Eating Cichlid Fish: Its Potential Role in Shaping Morphological Asymmetry PLoS ONE, 7 (9) DOI: 10.1371/journal.pone.0044670
Kusche, H., Lee, H., & Meyer, A. (2012). Mouth asymmetry in the textbook example of scale-eating cichlid fish is not a discrete dimorphism after all Proceedings of the Royal Society B: Biological Sciences, 279 (1748), 4715-4723 DOI: 10.1098/rspb.2012.2082
Takahashi, T., & Hori, M. (2008). Evidence of disassortative mating in a Tanganyikan cichlid fish and its role in the maintenance of intrapopulation dimorphism Biology Letters, 4 (5), 497-499 DOI: 10.1098/rsbl.2008.0244
Hori, M., Ochi, H., & Kohda, M. (2007). Inheritance Pattern of Lateral Dimorphism in Two Cichlids (a Scale Eater, Perissodus microlepis, and an Herbivore, Neolamprologus moorii) in Lake Tanganyika Zoological Science, 24 (5), 486-492 DOI: 10.2108/zsj.24.486
What is the largest living structure on Earth? No, it’s not the 2200 acre Armillaria ostoyae fungus in Oregon that we talked about previously. That is the largest single organism, but there is something bigger.
 |
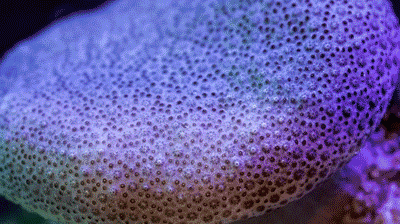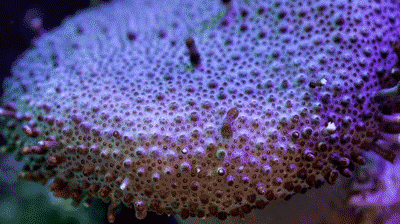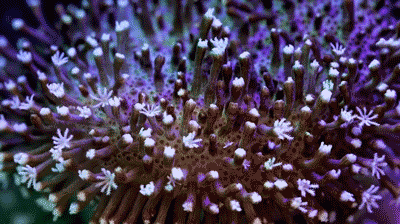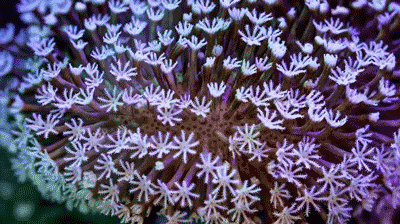
The Great Barrier Reef houses more species of coral than any other place on earth, more than 600 species call the reef home. You see how many shapes they can take. Does this mean they are asymmetric animals? |
This can add up quickly, because the Great Barrier Reef is more than 132,973 sq. miles (344,400 sq. km) in area. And if you still don’t believe it is a living structure, get this; it’s moving south! Climate change is warming the waters off the coast, and corals and tropical fish are moving south with the warmer temperature. Meanwhile, the northern edge recedes as the waters get too hot for corals.
Corals take all sorts of shapes (see picture above), and despite what we talked about last week, they don’t seem to be either bilaterally or radial symmetric. Sure, brain corals look radial, but most corals don’t have a repetitive shape. Are these asymmetric animals?
Nope - remember that the coral you see is the exoskeleton of the polyp, not the animal itself. Just because the apartment building isn’t symmetric, it doesn’t mean the animal is as well. Coral polyps are definitely radially symmetric, so our discussion of last week still holds.
Corals and other radial animals are exceptions, since 99% of animals are actually bilaterally symmetric. But there are exceptions with the bilateral animals as well. Some species that have been bilateral for millions of years then evolved a tweak to the system. Some part them became asymmetric in order to give them an advantage. Their stories are exceptional and we should explore some of them.
 |
Coral polyps are cnidarian animals. They live inside the calcium shells they produce, and it is the shells that seem asymmetric in many cases. But the polyps show radial symmetry. Each stalk and white “flower” is an individual polyp. |
Let’s start with the first animals that became bilaterally symmetric– the platyhelminthes, or flatworms. Most flatworms are small, just barely visible with the human eye, and most swim in the water. There are free-living versions and parasitic species and it is in the parasites that we find our first animal that has decided that completely bilateral isn’t necessary.
Polyopsithocotylea monogenea is a group of flatworms that live on and feed on fish gills. About 0.05-1 mm long, these platyhelminthes attach themselves to one of the gill ridges and take up residence there for life. The attachment they use is called a haptor, and they come in different shapes and sizes.
The oncomiracidium stage of the worm is directly after the egg stage and before it becomes an adult. This stage is completely bilaterally symmetric. But when the adult stage is reached and it’s time to settle down and starting feeding on some fish’s gills, they become asymmetric via their haptor attachment.
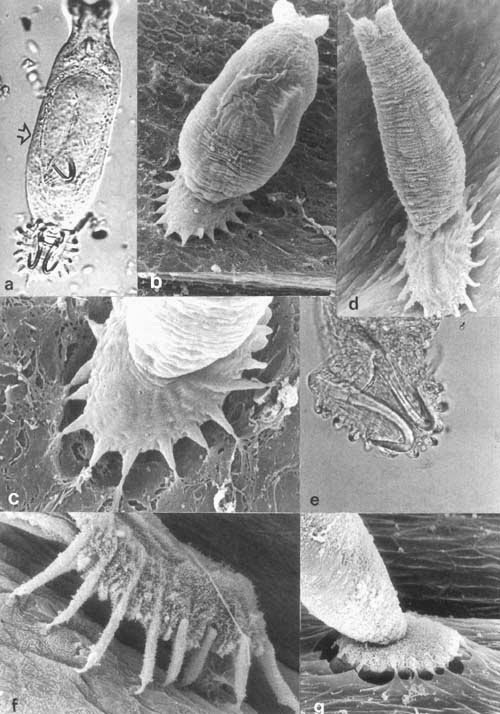 |
Opisthaptors, or just haptors, are the attachment organs for many parasites, including parasitic flatworms. They can use suckers or clamps and hooks in order to anchor the worm in its preferred habitat. You can see that these haptors maintain animal symmetry, but not all do. |
The initial haptor is usually located on the posterior end, on one side only. So much for bilateral symmetry. In some species this haptor has suckers, in others it has hard clamps, and yet other species have both.
As the adult grows, more haptors may develop, just where the animal touches the gill. Some may have 50 or more haptors arranged around their posterior, becoming more and more asymmetric. But there is a plan, they only grow where attachment is possible; some signal is generated by contact and this stimulates growth of more attachments.
Here we have a family of parasitic worms that aren’t symmetric living within a phylum of worms that were the first to be bilaterally symmetric - exceptional. But, take one step up the chain and you see the fish they live on. It just so happens that at least one group of fish parasitized by P. monogean worms are asymmetric themselves.
 |
These are the profiles of some monogean flatworms. The haptors of these parasites grown in odd places and destroy the bilateral symmetry of the animal. But it is necessary for the worm to attach to the gills of their prey fish. |
The cichlids are successful because different species have developed different feeding niches, and this is where we meet our asymmetric cichlids – they eat the scales off other fish! One genus, Perissodus, has at least six species that eat scales, all endemic to Lake Tangayiki (although there are also other scale eaters in other locations).
Scale ripping and eating is called lepidophagy (lepido = scale, and phagy = eat). Scales are an unexpectedly good source of nutrition. They chock full of protein and calcium phosphate, and their attachments are both cartilaginous, fatty, and come with some carbohydrate. Remember that the next time you order fish in a fancy restaurant. Tell the chef not to scale it – you’ll be quite the topic of conversation in the kitchen.
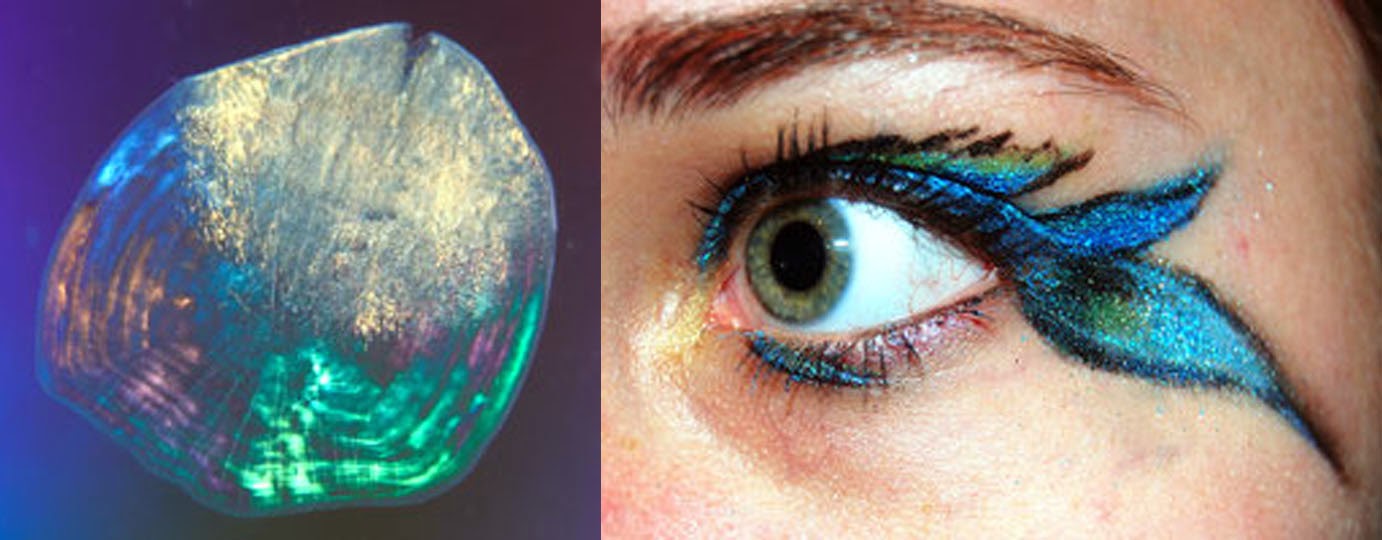 |
Fish scales have many uses. Some, especially from herrings, are used to make the pearlescent cosmetics that are sold today. Gals, your putting fish scales on your eyes and lips. Fish scales can also be turned into artificial bone, or they can be food for lepidophageous fish. |
But, you can imagine that the fish being unfrocked don’t appreciate it very much and fight back or swim away quickly. That doesn’t even take into account how hard it is to bite the scales off a swimming fish. Therefore, lepidophages must evolve anatomies and behaviors that give them a chance to succeed. Or perhaps it would be better to say, they acquired characteristics that made being a lepidophage an advantage.
 |
Here are two P. microlepis scale eating fish. One is right mouthed and the other is left mouthed. You can see how the way their mouth develops breaks their bilateral symmetry. The right-mouthed version (on the left) will only eat scales from the left side of fish. How is it that both versions can be maintained in a population? |
A 2012 video study showed that right-mouthed individuals almost always attack prey from the left, and their strikes are more powerful and successful when coming from the preferred side. One could ask, why are their both types? How did right- and left-mouthed individuals come to evolve and why are there still both types?
A different 2012 study shows that juveniles prefer one side or the other, even before their mouth bend has become pronounced, so it is a deeply penetrating characteristic, both heritable and perhaps partially acquired. There’s evolution and genetics at work here.
The prevailing model is that at any one time, right or left-mouthed individuals will predominate in a population. Let’s say that right now, right-mouthed feeders are the majority. The prey fish will learn to pay more attention to their left side, as this side is more vulnerable.
This makes it harder for right-mouthed individuals to feed, but easier for left-mouthed fish, because the prey fish ignore their right side relative to the left. In time, the right-mouthed individuals breed less well and the numbers in the population will shift. The left-mouthed feeders will become the majority. This is an example of negative frequency-dependent selection, where as a trait becomes more common it becomes less advantageous, and there is a balancing selection.
 |
The flu virus comes with one of many hemagglutin proteins and one of many neuraminidase proteins. If one genetic version is too successful, most people will develop immunity to it and it becomes less fit in the population of hosts. Its success is its downfall and a more rare version will rise up. This is negative frequency-dependent selection. |
However, a 2012 study throws this into question. They found equal distributions of right-and left mouthed individuals in five populations they studied. Also, left-mouthed and right-mouthed individuals mated with each other just as often as they mate with same-sided individuals (called disassortative mating). Perhaps the mouth bend is not a true dimorphism (di= two, and morph = shape).
Or, as a 2008 study suggests, negative frequency-dependent selection works best when there is disassortative mating. This may be necessary since a 2007 study showed that lefty:lefty matings give 2:1 lefty offspring, right:lefty mates give equal righty and lefty offspring, but righty:righty pairs ONLY give righty kids. Figure out the genetics of that. The authors proposed two possibilities – mendelian genetics with lefty being dominant and dominant homozygous being lethal, orcross-incompatibility that is predominant in lefty:lefty homozygotes, (meaning lefty homozygotes can’t mate successfully mate with the other types).
Today we have seen a swimming flatworm that feeds on some fish, and some fish that feed on the scales of other fish. And both of them achieve this only because they have adapted their bilateral symmetry to become just a bit asymmetric.
Next week, there are other animals that break symmetry to survive. Flatfish lay on their sides at the bottom of lakes and oceans, yet they still use binocular vision. How can that be?
For more information or classroom activities, see:
Great Barrier Reef –
Coral polyps –
Cichlids –
http://www.cichlid-forum.com/articles/lake_tanganyika_diverse.phphttp://www.news.leiden.edu/news/super-fast-evolution.html