Biological concepts – endoreplication, endocycling, endomitosis, decidualization, trophoblast, megakaryocyte
Last week we learned that polyploidy plays a role in cancer development and is the number one cause of spontaneous abortions in humans. Polyploidy is just no darn good.
 |
There’s alot to fret about once you hit the atmosphere. But take heart, you’ve already found a way to make a cancer-like pathway work for you. Don’t worry about the details, you’ll get it all in Biology class. |
For example, osteoclasts (osteo = bone, and clast = to break) form from the fusion of two or more precursor cells. Since each precursor cell has its own nucleus with a 2n set of chromosomes (n=23 for humans), the fused cell may have 4n, 6n, 8n, or more chromosomes, in one or more nuclei. New evidence shows that not only can they fuse, but they can also fission to form more osteoclasts when needed. This had not even been hinted at before.
Osteoclasts eat bone; you are forever tearing down bone and replacing it with new bone. If you lift weights and build bigger muscles, you need bigger bones onto which you can attach your now stupendous guns. About every ten years or so, you have an entirely new skeleton!
Polyploid cells can be formed when diploid cells fuse, but it is more interesting when they are formed by the processes of endoreplication (endo = within). Normally, most cells just hum along, growing (G1), then replicating their DNA (S), then growing some more (G2), and finally dividing into two daughter cells by mitosis(M). The two new cells then repeat the process. This is called the cell cycle, and is abbreviated as G1, S, G2, M.
The mitosis portion of the cell cycle itself has several parts that we all learned in biology class – shout them out with me - prophase, metaphase, anaphase, and telophase! At the end of telophase, the two daughter cells finally decide they can’t be roommates any longer, and they divide up their belongings.
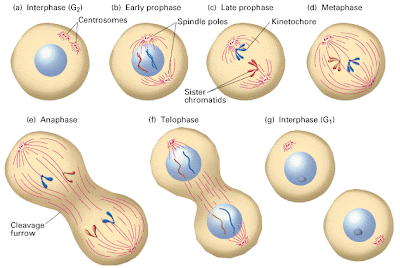 |
The phases of mitosis finish up by dividing the cytoplasm and nucleoplasm. You can see that the cytokinesis starts first, with the appearance of the cleavage furrow, but karyokinesis is completed before cytokinesis is done. Therefore, you can’t have complete cytokinesis with defective or incomplete karyokinesis. |
In endoreplication, one or both of these processes is turned off, so the two daughter cells continue to share a room, but now the room has twice as much DNA (4n instead of 2n). The cell skips at least a portion of M phase, and the cell cycle becomes G1, S, G2 ----G1, S, G2, etc. It may occur just once, producing a tetraploid cell, or it may occur several times, forming huge cells with 32n or more chromosome sets.
If the cell skips mitosis all together, the process is called endocyling. In this case, the chromatids don’t separate in anaphase, and you end up with chromatids that remain stuck together at their centromeres. If they replicate again in the next S phase, you end up with an octopus-looking chromosome with several arms sticking out – called a polytene chromosome.
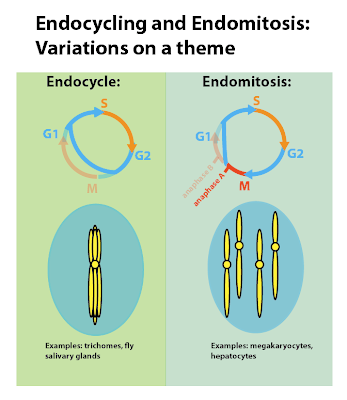 |
The left side of the cartoon shows endocycling. Skipping mitosis altogether keeps the chromatids connected and forms polytene chromosomes. Endomitosis is on the right, where the cell goes through part of mitosis, then skips the part where the two cells separate, either by skipping cytokinesis alone or karyokinesis and cytokinesis. |
Polytene chromosomes occur naturally in some animals, like the huge (1 mm) chromosomes in the salivary glands of larval fruit flies (Drosophila melanogaster). They benefit the fruit fly larva in that the cells can produce more proteins from the many copies of the genes. This allows the fruit fly larva to make enough of the proteins that are important in forming the pupal case when it undergoes metamorphosis. All due to endocycling and polyploid formation.
On the other hand, if a cell starts through mitosis and separates its chromatids, AND THEN decides to not divide, this is called endomitosis. Cells that have undergone endomitosis have many sets of chromosomes. Endomitosis without cytokinesis results in large cells with multiple diploid nuclei because karyokinesis separated the nuclei. Endomitosis without karyokinesis and cytokinesis results in large cells with a single polyploid nucleus. You can see that polyploidy would need to be highly regulated to keep it from getting out of control.
So how is that polypoloidy is crucial for our survival? It turns out that that some specialized cells of the embryo undergo polyploidization as the embryo implants into the wall of the uterus.
The embryo has an outer layer of cells called the trophoblast; these cells become the placenta, attach the embryo to the uterine wall, and create the blood vessel connection between mama and junior. The trophoblast is the first set of cells to differentiate in the embryo and they become several different types of trophoblasts.
One type in particular, the extravillous cytotrophoblasts (ECTs), spread out from the developing placenta and burrow into the uterine wall. This creates the tight attachment between mom and embryo. The ECTs also send out hormones to rearrange the mother’s blood vessels, forming the umbilical cord and vessels. This is how the growing baby gets all its nourishment until delivery.
ECTs have been studied most in rodents; they weren’t recognized in humans until just recently. However, a late 2012 study has shown that ECTs are released from the placenta and can be studied by collecting them at the cervix. The cells were sufficient to determine the sex of the child after only 5 weeks of gestation, and were generally of 4n-8n ploidy. Interestingly, female fetuses tended to form ECTs at a rate almost 7x higher than male fetuses – you’re guess is as good as mine as to why that might be.
 |
In panel A there is a bunch of abbreviations. VT is the villous trophoblasts that make the connection to the decidua (DD). In the black box which is enlarged in panel B, you can see the extravillous cytotrophoblast (EVT here) cells invading the decidua. Both the EVT and the decidua are polyploid. |
The reason that cells of the decidua must be polyploid is unknown, but the fact that polyploidization begins at the point of implantation and spreads to a greater part of the uterus tells you that they are necessary. A new study points to a few possiblereasons. Comparing polyploid decidua to non-polyploid decidua showed that many genes were up-regulated or down-regulated.
The up-regulated genes had to do with metabolism, especially the mitochondrial energy production. On the other hand, down-regulated genes had to do with apopotosis and immune function. These results suggest that polyploidization of the decidua is meant to increase cell functions for the benefit of the embryo, and this takes energy (so more mitochondrial function), while at the same time making sure the cells survive to support the fetus until delivery (reduced apoptosis gene function) and protection of the fetus from the mother’s immune system (the baby is a foreign body after all).
So baby has polyploid cells that mediate joining with the mother, and mom has polyploid cells that also work in the formation of the link between the two. Everyone has to bring polyploidy to the party, or ain’t nobody getting born!
However, polyploidy in fetal development is only part of the story. You don’t abandon polyploid cells altogether once you are born or give birth. All of us have polyploid cells in our bodies right now. Take megakaryocytes for instance.
When you cut yourself, or there is a leak in a blood vessel, platelets arrive to help close the hole and stop the bleeding. Platelets are of irregular shape and are sticky, so they tend to get stuck along the edges of broken blood vessels. Then other things stick to them, a few dozen enzymatic reactions take place with myriad proteins, you form a clot (called a thrombus in the medical world).
 |
I have described how platelets are important for coagulation. This cartoon shows a break in the cell on the bottom, and ALL THE STUFF that has to happen to forma thrombus (clot). Platelets are central, but they are certainly not all the story. |
Hepatocytes(liver cells), smooth muscle cells in blood vessels, heart muscle cells – these can all be polyploid. In hepatocytes, polyploidization occurs in cells that are done dividing and specializing (terminally differentiated) and are now just doing their job. Fetal and newborn liver cells are exclusively diploid, but 30-40% of adult hepatocytes are polyploid.
Polyploidy may be a way to increase liver metabolism and function without going through cell division. Or it may help to protect the cell from the effects of individual mutations. Since the liver is involved in breaking down toxins, it’s a good guess that some genes will mutate. Having extra copies around would prevent a mutation from inhibiting cell function. One mutated gene can be compensated for by an additional normal gene.
On the other hand, smooth muscle cells seem to undergo polyploidization as a prerequisite to senescence; they are aged and they just stop working. This interesting, since we said last week that cancer cells are more likely escape therapy induced senescence by becoming polyploid. Once again, biology can turn the ordinary on its head.
We have discussed the appearance of a polyploid mammal and crucial sets of polyploid cells in humans. These are the exceptions in higher vertebrates. But in other organisms, polyploidy is a key to evolution. Next time we’ll talk about the exceptional role of polyploidy in the development of plants.
For more information or classroom activities, see:
Osteoclasts and bone remodeling –
Endoreplication –
http://www.news-medical.net/news/20111031/Researchers-shed-light-on-inner-workings-of-endocycle.aspx
Trophoblast and decidualization–
Megakaryocytes –