Biology concepts – parthenogenesis, gynogenesis, kleptogenesis, sperm-dependent parthenogenesis, pseudogamy, Muller’s ratchet
Last week we introduced the idea that species can be (facultative) or must be (obligate) parthenogenic. Both facultative and obligate species are diverse, interesting, and full of exceptions – what a surprise.
 |
The pea aphid is a wonder of biology. Here, you see a winged male with offspring nearby. It is hard to tell if these are clonal offspring, but they are likely to be found in the Fall, as winged males are produced from late summer eggs. This is so they can fly to new food if necessary, before mating and the females laying eggs that will overwinter. |
Overwintered eggs hatch in the spring and become wingless females. These individuals immediately begin to give birth to clones of themselves,apomictic, thelytokic, parthenogens. These are all females, due to the sex determination system that aphids use, the XX/XO system. When diploid develop, they double their haploid chromosomes, so all are XX females.
The parthenogenic females reproduce quickly, giving birth to dozens of females over a period of just days. These females immediately begin to give birth to more clonal females. The reason it can be so fast is that the females are born pregnant! The process is called telescoping generations, because there is less and less time between birth and birth. This is one form of paedogenesis (paedo = child), reproduction by sexually immature forms.
 |
The life cycle of the pea aphid is complicated, having both sexual and asexual components. In the spring to summer, females will produce off spring by parthenogenesis. In the late summer and Fall, the parthenogenic females will mate with males and lay eggs that will hatch in Fall and later eggs that will hatch the next spring. |
The result is that millions of offspring can be produced from a single female in the spring (although they live only about 10-40 days). A comparison is warranted. If all the offspring from a female lasted an entire summer and they were lined up in a single line, they could circle the Earth more than four times! Maybe there is something to this parthenogenesis.
Bees are also facultative parthenogens, but with a different twist. Bees are haplodiploid, meaning that all the males develop from unfertilized haploid eggs, while the females come from fertilized eggs. Even the sterile female workers are the result of fertilization. The twist comes when in some species, the queen dies without an heir. In this case, some of the sterile worker bees can start to lay eggs. It is a futile effort though, they produce only males because they are sterile and have not mated. The hive dies out anyway.
The exception to this unfortunate affair is one species of South African bee, Apis melifera capensis, who can repopulate by hiring a new queen. The female workers of this species will fight it out when a queen dies, and some will start to produce diploid eggs to produce a new queen by parthenogenesis. She will be a clone of a worker, but she will mate with a male and introduce more genetic diversity into the hive.
 |
Some species of whiptail lizards are females only – no males at all. But they need the stimulation of feigned mating to start development of the unfertilized eggs. So females who have just laid eggs act as males and perform male behaviors. Females that are acted on by these “male fakers” are more likely to lay eggs and have the young survive. |
Other parthenogenic species need more help to jump-start the egg development. Many species require sperm in order to stimulate egg development. The sperm does not contribute any DNA to the embryo, but it contains a chemical, hormonal, or physical property that makes the egg develop into a whole animal.
If many obligate parthenogens are strictly female, where does the sperm come from? A male of a closely related species usually does the honor, but it doesn’t really matter, since the DNA is not incorporated into the egg. This process has many names, and they all mean pretty much the same thing - sperm-dependent parthenogenesis, kleptogenesis, pseudogamy, gynogenesis– more names than those two fellas on “Psych” (when are they going to bring that show back?).
 |
The amazon molly doesn’t live in the Amazon River. It was named for the Amazon warriors of Greek mythology, an all female warrior society. The amazon molly is an all female species that reproduces by gynogenesis. They mate with a closely related male, but do not incorporate his DNA into the developing embryo. The sperm is needed to stimulate egg development. |
Many times, parthenogenesis is an animal’s only choice, but there are definite advantages to this mode of asexual reproduction. One, the offspring are clones, produced under a certain set of environmental conditions. Since the conditions were good enough to let the mother survive and reproduce. That means that offspring exactly like her should thrive in those conditions too. Little effort – maximum effect.
Two, we talked last week how rapid reproduction by parthenogenesis can help komodoscolonize new territory quickly, much faster than they could by sexual reproduction alone. And three, parthenogenesis doesn’t waste community resources and energy on animals that don’t give birth – males. I don’t think I like this advantage.
But there are also definite disadvantages to parthenogenesis. One disadvantage is that the very clonality that helps them in steady state conditions is a hindrance if the environment changes. Genetic diversity is important for adaptation, but parthenogenesis offers no chance for genetic diversity.
Another potential disadvantage to parthenogenesis is the loss of traits that are needed for sex, like mating behaviors, mating calls, etc. An example is a facultatively parthenogenic fruit fly. In 1961 they were separated from males and raised separately. Ten years later they were reintroduced to males. Only some mated, but they still had the genes that controlled mating behaviors. I 1981 they were reintroduced again, and none of the females participated in the mating behaviors; they had been lost completely.
 |
Muller’s ratchet has more to say than just that unused genes will drift. In terms of becoming parthenogenic, it does surmise that genes that have to do with sexual reproduction will mutate at a higher rate. However, it also states that there will be deleterious mutations in asexual organisms, resulting in a drop off in births. As such, the ratchet is a commonly held argument for why sexual reproduction is so evolutionarily important. |
One last disadvantage - parthenogenic species seem to last only about 100,000 years on average, probably due to the lack of genetic diversity. However, some salamanders have been gynogenic for 1 million years, suggests that they have had a few indiscriminate fertilizations along the way that have introduced new DNA, about 1 in a million births. Some orbatid mites (1 mm soil mites that help recycle dead material) have been parthenogenic for 100 million years!
Even though species have been parthenogenic for millions of years, it is only in the last few decades that we have really learned anything about these behaviors. Now that we have some knowledge, it seems time to put it to use.
For instance, human eggs can now be induced to develop in the absence of sperm. Before release, pre-eggs are frozen in time in metaphase II stage of meiosis. This means that they are still diploid, it isn’t until anaphase and telophase that the chromatids are pulled apart and the eggs become haploid.
In this stage, if you prick the eggs with a needle on their membrane, or treat them with some chemicals, or apply a mild electric shock, it seems to bring the same response that penetration of a sperm head does. This triggers the initial stages of development in the egg (blastocyst), regardless of the fact that it doesn’t have dad’s DNA.
Under these conditions in the lab, the eggs will develop to the 500-1000 cell stage, and then they will die out. Remember that they do not have the paternally imprinted genes available to them, so they can never become a full-fledged embryo.
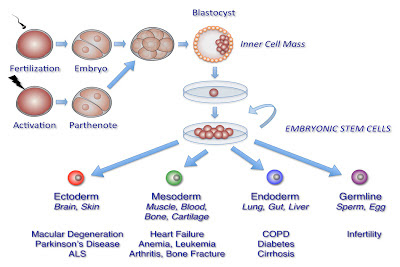 |
Human stem cells are produced by teasing out the cells of a blastocyst and growing them separately. Then you can treat them with different growth hormones and make them into different types of cells. One way to get the blastocyst cells is from fertilized eggs. But to avoid those ethical headaches, now scientists often stimulate the egg to develop parthogenetically, and then harvest the stems cells. |
For instance, one 2012 study showed that hpESC’s could be used to generate mesenchymal stem cells, that had the ability to differentiate into several different type of cells, include bone making cells and fat making cells. They compared the hpESC’s to stem cells generated from embryos and found they expressed very similar marker proteins. Because they are homozygous for immune markers, it is hoped that hpESC’s will be good replacement cells in tissue therapies.
Next week – birds can undergo parthenogenesis, but it is usually not a happy ending, unless you like omelets.
For more information or classroom activities, see:
Sperm-dependent parthenogenesis –
Mueller’s ratchet –
Human parthenogenic embryonic stem cells –