Biology concepts – homeostasis, neuroendocrine system, hormone, pituitary, hypothalamus
 |
Let’s face it, everything I know about computers I learned from Tron and Tron Legacy. What I learned most from the sequel is that we still can’t make a decent avatar for Jeff Bridges. But I did learn about the CPU and programs and users. I like to think there are small motorcycle races going on in my laptop while I write. It makes it more interactive. |
Basically, a computer is like any other tool, its worth depends on how it is used. But its a very complex tool with many parts that are easily broken or can wear out. A computer is like your brain in some ways; let’s look at one specific system in your body that can be likened to part of your computer.
The central processing unit (CPU) of your computer controls most of what your machine can do, but it’s functions fall into two broad categories – 1) calculations and information storage, and 2) monitoring the machine itself. The CPU has programs to manipulate information and present information in different forms, but it also has sensory and feedback systems to keep the machine working at optimum levels.
It is this second function that we’re interested in today. Let’s say you use your laptop for an extended time and it starts to heat up. Sooner or later you'll hear the fan turn on; this is your computer’s attempt to maintain the temperature within a small window of values so that function is maintained and there is no damage to the machine. Your body does the same thing.
Another example – you let your computer sit for a while without doing anything. Sooner or later the screen will go black and the machine will go into a sleep mode to save energy and not burn out the screen. Once you start to use it again, it will draw more power; if you stream a movie or 100 videos of cats playing the piano, it will draw a lot more energy and the battery will wear down faster.
 |
Homeostasis is a little like riding a Segway. If you stand perfectly still and upright, nothing moves; all the sensors are working but they don’t have to make any adjustments. If you lean forward, the gyroscopes recognize it and move the Segway forward so that you stay balanced; the same thing if you lean backwards. Everything works to keep your body systems in balance even when, especially when, something changes in the system. |
In your body, the brain is your CPU. You’re cerebral cortex controls your higher functions, but each hemisphere has older parts that help maintain the body systems the hypothalamus and hippocampus (so do you have dual processors?). The peripherals that help control your body and keep things on normal are the endocrine glands and the neuroendocrine cells. Together, they’re called the neuroendocrine system and they maintain homeostasis (homeo = like and stasis = standing still).
Homeostasis is one of the characteristics of life. To be alive, an organism needs a system to resistant changes in itself when there are short-term changes in its environment. Like a computer overheating and turning on the fan, your body is constantly sensing internal temperature and turning on and off discrete systems to maintain a constant 98.6˚F.
You have homeostatic systems that control reproduction, temperature, energy consumption and production, hunger, sleep/wake cycles, basal metabolic rate, osmolarity, drinking, and blood pressure. Some of the sensory systems run straight to the brain via hardwiring (another computer analogy). These are the neurons of the peripheral and central nervous systems.
The outputs might be neural, but many times they aren’t. Imagine trying to send a neuron to each cell that needs to get a chemical message (via neurotransmitters released at the ends of the neurons). That would be very cumbersome to maintain and would require trillions more neurons.
 |
“To whom it may concern,” is the introduction to every hormone message sent throughout your body. Like bulk e-mails, the hormones contact every cell via the blood system, but only those that can and need to respond (have the right receptors) will read the message and do something about it. |
Hormones are the hallmark of the endocrine (endo = within, and crine = sift) system. When released into the bloodstream, they travel to every part of the body so that all cells have the opportunity to respond. Not all will, and you wouldn’t want them to. Specific cells and organs have the ability to respond to specific signals because they have the right hormone receptors.
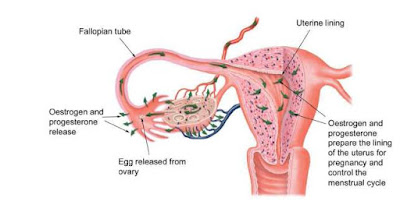
There are eight endocrine glands in a human body. The adrenal glands (2 of them), the parathyroid glands (4), the thyroid gland (1), the pineal gland, the pancreas, the ovaries (2) or testes (2), the hypothalamus, and the pituitary gland. They can be stimulated to release hormones by either neurons, other hormones, or other chemicals. Besides these well known glands, a few tissues will release hormones in specific situations. The placenta will release progesterone and estrogen, and the stomach can release gastrin to stimulate gastric juice (acid) and ghrelin to stimulate hunger.
In addition to endocrine glands, there are also some cells that can be directly stimulated by neurons to release hormones into the blood. These are the neuroendocrine cells. There’s a lot more of them you think, and they’re just about everywhere in your body.
 |
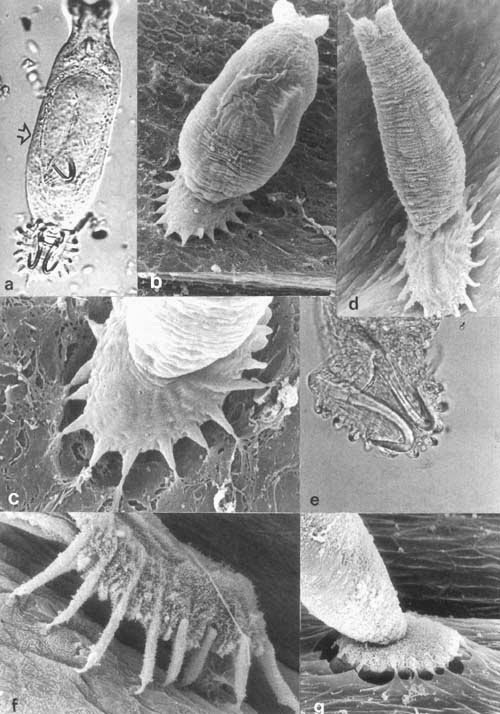
The upper image shows the PNEC’s in your respiratory tract (NE). They can be alone or be grouped into neuro- endocrine bodies (NEBs). A 2008 study, and several before it, has correlated a hyperplasia and hypertrophy of PNECs with Sudden Infant Death Syndrome. Too many and your respiratory control may get thrown out of whack; it may just stop working. It might be good to monitor the PNEC system in youngsters to try and predict susceptibility to SIDS. |
For one example, there are pulmonary neuroendocrine cells (PNEC) in every part of your respiratory tract, from your nose to your alveoli. While they seem to act like neurons in many respects, they are derived from epithelial tissue, not the neural crest tissue that all neurons come from.
Each PNEC spans the distance from basement membrane to the air conducting space. On the luminal side, they have microvilli that stick out into the lumen and sample the air as it passes. On the basal side (bottom) they communicate with neurons. Just what are they doing there?
They have several functions, including regulating the development of the respiratory system in the fetus. They also regulate the function of the respiratory cells by sensing oxygen levels, controlling the muscular tone of the bronchi, regulating pulmonary blood flow and modulating immune responses. The origin and function of PNEC’s is reviewed in a 2012 paper, which also highlights a problem with them – when PNEC’s go bad, they cause a deadly small cell cancer.
In response to what the cell senses, it will release a variety of chemicals, many of which can act as neurotransmitters – hence the reason they are considered neuroendocrine cells. But the PNEC’s may have another function, one hinted at in a 2014 study. The results of these experiments indicated that PNEC’s can detect chemicals in the air as it passes over their microvilli – they quite literally can smell the air, as their microvilli were found to have olfactory (smell) receptors, just like in your nose!
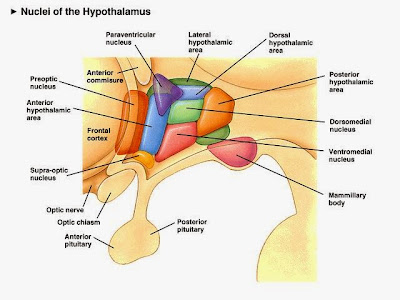 |
The hypothalamus is part of the diencephalon, a part of the brain older than the cerebral cortex, but not as old as the thalamus. The different nuclei each function in different systems; the lateral nucleus senses hormones and regulates thirst and hunger, the dorsomedial nucleus regulates BP and HR. Several nuclei are involved in making releasing hormones, ADH and oxytocin. |
So what controls much of this neuroendocrine system? Your brain of course. Well, a small part of your brain that's pretty old in terms of evolution (even primitive hagfish have a pituitary). The hypothalamus senses many of the inputs that tell your brain just how well your body is maintaining homeostasis. If something goes astray, or if there is a change in your environment that forces a change in your body chemistry, the hypothalamus will then spring into action (O.K., it’s always working, it will just work harder).
The hypothalamushas a neuroendocrine relationship with the pituitary gland (better named the hypophysis, where hypo = under and physis = growth – it is a growth under the brain). The hypophysis comes in two parts. Both parts release hormones, but where those hormones come from and the sources of the tissues of the two pituitary parts are very different.
The anterior pituitary isn’t part of the brain at all. The tissue for the adenohypophysis(anterior pituitary, adeno = secreting) comes from the roof of the embryonic mouth. It is epithelial in origin and has cells that produce hormones in response to signals from the hypothalamus (also hormones). Here’s how it happens.
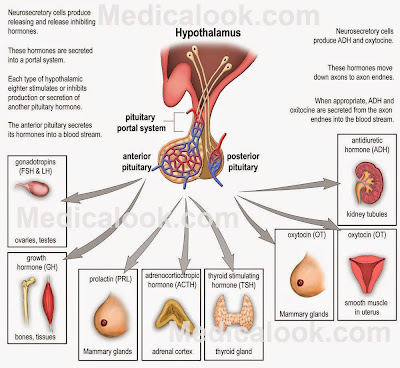 |
The pituitary gland, or hypophysis, has two small portal systems that carry blood from one place to another instead of to a particular tissue and then back to the heart. The anterior system carries releasing hormones from the hypothalamus to the ant. pituitary and then carries stimulating hormones from the ant. pituitary to the rest of the body. The posterior system moves hormones from the hypothalamus directly to the rest of the body. Image credit to Medicalook. |
Well, it gets worse. There are six releasing hormones from the hypothalamus that stimulate production and release of seven hormones from the adenohypophysis that then act on at least 20 endocrine glands and neuroendocrine cell types. Hypothalamus to adenohypohysis to endocrine gland – it’s called an axis, and there are several of them.
On the other hand, the posterior pituitary (neurohypohysis) is derived from brain tissue. Instead of the neurons of the hypothalamus producing releasing hormones that then act on the neurohypophysis, the hypothalamic neurons project right into the posterior pituitary where they deposit their hormones (oxytocin and antidiuretic hormone). The neurohypophysis doesn’t make any hormones itself, it just stores what the hypothalamic neurons produce and then releases them to the circulatory system.
That’s a heck of an exception- part of your brain isn’t actually part of your brain. Cells from your mouth control most functions in your body! Some people I know have mouths that completely ignore their brains! Next week, let’s talk more about the neuroendocrine system. Your thyroid gland size correlates to which hand you use to write – say what?
For more information or classroom activities, see:
Homeostasis –
Neuroendocrine system –
Hypothalamus –
Pituitary –