Biology concepts – hydrogenosome, FeS cluster protein, loricifera, erythrocyte
 |
More than one way to skin a cat seems to be a newer version of the old British saying, “there are more ways to kill a cat than by choking it with cream.” Mark Twain was one of the first to use the cat skinning version, in his classic A Connecticut Yankee in King Arthur’s Court. |
The oldChinese proverb says, “There are many paths to the top of the mountain, but the view is always the same.” Put somewhat less delicately, “There’s more than one way to skin a cat.” Who wants to skin a cat? I think there is something to be said for the wisdom gained in 4000 years of culture, to say nothing of the ability to say it better.
In biology, this is particularly relevant; organisms have found different ways to do the same things, and different ways to do different things, but the end goal is always the same – live long enough to reproduce and the more offspring the better.
Last week we talked about how some organisms have degraded their mitochondria into mitosomes, and how they get along fine just using glycolysis and fermentation for energy (and maybe some arginine dihydrolase action). But there is another mitochondrial remnant in some other species of anaerobic eukaryotes called the hydrogenosome, and it works more like a mitochondrion than does the mitosome.
 |
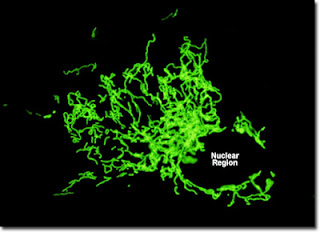
Here is the T. vaginalis protist. The blue probe binds to DNA (just one nucleus for this guy) and the yellow probe binds to a hydrogenosome protein. The strands at the top are the flagella it uses to move, not its hair. |
Trichonomas vaginalisis a eukaryotic amitochondriate, and therefore is an anaerobic (without oxygen) protozoan. Unlike many protozoans, T. vaginalis does not have an environmentally resistant form (something that can live outside the host for a prolonged time – often called a cyst). It is transmitted directly from host to host, in this case sexually. Trichomoniasis is the most common curable sexually transmitted disease, but 70% of cases have no symptoms (asymptomatic). This is unfortunate because T. vaginalis infection can predispose to HIV infection and even cervical cancer. Having symptoms initially might prevent some of the later tragedies.
Unlike the mitosome containing protists, T. vaginalis does use its mitochondrial remnant (hydrogenosome) to make ATP. The hydrogenosome was discovered much earlier than the mitosome, although they have the same origin and general morphology. Because of this difference in timing, amitochondrial organisms with hydrogenosomes are called type II amitochondriates. Type I’s were the organisms that presumably didn’t have any mitochondrial-like organelle (and were seen first), like the Giardia and E. histolytica that we now know have mitosomes.
Pyruvate generated by glycolysis enters the hydrogenosomes just like it does in mitochondria. The Krebs cycle would be next for aerobic organisms, but in the hydrogenosome, iron-containing enzymes convert the pyruvate into an intermediate that has CoA (coenzyme A) bound to it. When this CoA is removed, energy is released, and this energy is used to convert ADP to ATP.
Because ATP production occurs at the level of substrate (a molecule being chemically changed, in this case by an enzyme), it is called substrate level phosphorylation. This is in contrast to the use of oxygen and the electron transport chain of proteins to produce ATP through the proton gradient (oxidative phosphorylation). One of the byproducts of the pathway is hydrogen, hence the name of the organelle.
In terms of energy production, the pyruvate:ferredoxin oxido-reductase (the iron/sulfate-containing enzyme in hydrogenosomes, often abbreviated as FeS cluster enzymes) pathway is about as efficient as the arginine dihydrolase pathway(ADH) in some mitosome-containing organisms. However, T. vaginalis also contains the ADH pathway, so it comes out ahead of Giardia in terms of energy production.
While the hydrogenosome has some activity in energy production via the FeS-protein mediated metabolism of pyruvate with production of ATP, the mitosome seems to be limited to the assembly of the FeS clusters only. A study of the proteins of the mitosome show the parts are there to make the FeS clusters, but that there are not the enzymes needed to break down pyruvate and produce ATP.
 |
A study trying to quantify the amount of methane gas produced by cows was carried out recently in Argentina. The method involved a big backpack and a delicately placed rubber hose. At some point, scientist A approached scientist B and said, I’ve got a great idea….” |
Other hydrogenosome-containing organisms include the anaerobic unicellular fungus, Neocallimastix frontalis (it lives in the guts of rumen animals like cows). N. frontalis byproducts are used by gut methanogens (methane-producing bacteria) and therefore contributes to the generous amount of gas produced by cows. Many estimates name dairy and beef cattle flatulence as a bigger source of greenhouse gases than automobiles!
Another hydrogenosome-containing protozoan is Nyctotherus ovalis. It lives in the GI tract of cockroaches, and efficiently works with an archaeal bacterium that uses the hydrogen that the hydrogenosomes release. Just one more reason that cockroaches will outlast us all. The fact that some fungi and some protozoans have hydrogenosomes indicates that this organelle has evolved independently from mitochondria at least three different times in history – they must be a good idea.
Even with the exception of anaerobic protists and fungi, it was believed until just recently that at least all multicellular eukaryotic (metazoan) organisms depended aerobic respiration for energy production. However, there are even metazoan exceptions. A 2010 study of the bottom of the Mediterranean Sea found three different animals that survive without using oxygen and therefore don’t have mitochondria.
The deepest basin of the Med, near Greece, is nearly anoxic (an environment without oxygen). In the muds of this basin were found three loriciferan(lorici = corsette and fera = bearing, so organisms with a sort of girdle) species that live in this area all the time. Other animals can survive in an anoxic environment for a while, but they don’t call it home.
 |
Loriciferans weren’t even discovered until 1983. Now we have some that live as anaerobes. Most species of this phylum live in the deep waters, but only a few are obligate anaerobes, meaning they can only perform anaerobic respiration. Oxygen can be damaging, it likes to scavenge electrons, I wonder if it is toxic to the loriciferans. |
These new loriciferans have hydrogenosomes instead of mitochondria, and produce ATP in the same ways as T. vaginalisand the other anaerobic eukaryotes. This is a completely new door being opened in biology, because the multicellular animals evolved after Earth turned from an anoxic environment to a place where oxygen was plentiful. It seems that even some of the more advanced organisms don’t have a problem reverting to more ancient systems if they find themselves in a place where they need it.
Would you believe that some of your cells might not have mitochondria? Well, about 26 trillion of your cells (if you’re an adult male) are amitochondrial – your red blood cells. That’s right; the erythrocytes that deliver oxygen to your cells in order so they can make ATP in their mitochondria don’t have any mitochondria of their own! In an attempt to carry as much oxygen as possible (bound to a big molecule called hemoglobin) your red blood cells have evicted their mitochondria.
This is probably a good idea, since making energy in the erythrocytes would use up the oxygen they are supposed to deliver to other cells. Instead, they act more like prokaryotes, and carry out glycolysis and lactic acid fermentation in their cytoplasm for the energy they need. To gain more room for hemoglobin, the RBCs have also done away with their nucleus. They have no way to produce more proteins or repair themselves, so they work as long as they can and then they are replaced.
Old erythrocytes are phagocytosed (eaten) by macrophages in the spleen and liver and are destroyed. New RBCs (about 2 million per second) are produced in your bone marrow. The spleen also acts as a reservoir for blood cells, a ready supply for when you need them, but you can get along without it, you are just more susceptible to infections, since the spleen houses many white blood cells just waiting to recognize a pathogen that needs to be taught a lesson.
 |
Human red blood cells (left) are round and biconcave, but the camel RBCs are oval. You can see why so many people believe they have a nucleus, but what you are seeing is their biconcave side staining darker. The large cell in the middle is an immune cell. |
Anucleate (a = without, and nucleate = pertaining to a nucleus) erythrocytes are the norm for mammals. Many people think that camels are the exception, that they have nucleated RBCs, but this is not so. But they do have ovoid RBCs. When they run low on water, camels can remove water from their blood and use it in their cells. This leaves their blood thicker and harder to push through the small capillaries. Round RBCs would be impossible to squeeze through when the blood is viscous, so the camel has evolved RBCs that are longer in one direction and smaller in the other, to help blood flow in times of dehydration.
On the other hand, almost all non-mammalian vertebrates do have erythrocytes that do have nuclei. The only exceptions are a few salamander species that have some anucleate erythrocytes. For example, 95% of the Batrachoseps attenuatus salamander’s RBCs are anucleate. There is also the pearlside fish which is known to have non-nucleated red blood cells.
However, the crocodile icefish is even a bigger exception; it is the only vertebrate animal that has gotten rid of its RBCs altogether. This species lives in cold, highly oxygenated waters. The oxygen it needs just travels in the blood as a dissolved gas and is carried to every cell. These fish have even lost the DNA for making hemoglobin – now that is efficiency!
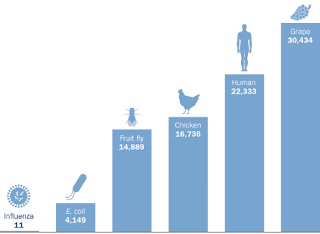 |
Given our apparent complexity, it is amazing just how few genes humans have; the grape has almost 30% more. The chicken doesn’t have many fewer than us, and we don’t have to worry about laying eggs. What is more amazing is that nine years after the completion of the human genome project, we still aren't exactly sure how many genes we have. |
Or is it?We have recently discovered that the majority of proteins have more than one function. Scientists gave this idea more thought when the results of the human genome project started to role in and we discovered far fewer genes than we expected. It is now accepted that humans have about 22,000 genes, not even as many as the grape, which has 31,000. Even the lowly fruit fly has 15,000 genes! How do we get so many functions out of so few gene products? Multitasking!
Take hemoglobin for example, it doesn’t just carry oxygen in the blood. It also acts as an antioxidant in several types of immune cells, and in certain neurons. It is a regulator of iron uptake and metabolism, since it carries iron at its core. It destroys nitric oxide, which is one reason why the little blue pill doesn’t work forever. You have to wonder what else the crocodile icefish has lost by giving up its hemoglobin and how it has made up for these losses. One change probably requires many more to be made as well.
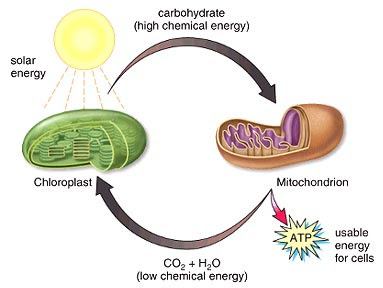
We have seen how some organisms get along without mitochondria. What about the other end of the energy equation? Plants can make their own carbohydrate in the chloroplast – but is that what makes it a plant? Let’s look at this next time.
Roberto Danovaro, Antonio Dell'Anno1, Antonio Pusceddu, Cristina Gambi1, Iben Heiner and Reinhardt Møbjerg, & Kristensen (2010). The first metazoa living in permanently anoxic conditions. BMC Biology DOI: 10.1186/1741-7007-8-30
For more information or classroom activities on hydrogenosome, FeS cluster protein, loricifera, erythrocyte, see:
Hydrogenosome –
FeS cluster protein –
Loricifera –
Erythrocytes –